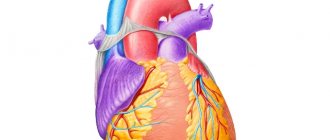
Physiological hypertrophy[edit | edit code]
The following signs are typical for physiological cardiac hypertrophy in athletes: 1) certain amplitude characteristics of the teeth of the QRS complex; 2) lack of QRS expansion; 3) normal time of local electronegativity; 4) absence of disturbances in depolarization of mixing downwards of the S-T segment, deformations or inversion of the T wave; 5) increase in heart volume over 11 ml/kg.
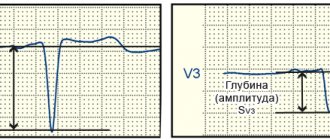
The ECG shows signs of hypertrophy predominantly of the left ventricle. The most sensitive electrocardiographic indicators of physiological left ventricular hypertrophy in athletes, in order of decreasing accuracy, are: Rv5-6 > 33 mm, SV1 + Rv5-6 > 53 mm, sum of QRS amplitudes in leads V2, V5, aVF > 93 mm, Ravf 23 mm, Rmax + Smax in precordial leads > 55 mm.
The most sensitive electrocardiographic indicators of physiological right ventricular hypertrophy in athletes, in order of decreasing accuracy, are: Rv1 + Sv5 > 11 mm, SV5-6 > 7 mm, Rv1 > 7 mm. Electrocardiographic signs of right ventricular hypertrophy are less reliable than signs of left ventricular hypertrophy. The combination of several signs of hypertrophy on the ECG allows us to speak with greater confidence about its presence.
It is possible to use separate indexes. Thus, with left ventricular hypertrophy, the Sokolov-Lyon index (mV) and the total voltage of the R waves increases in both standard and left chest leads. Moreover, these signs are more pronounced in men.
When analyzing an ECG, it is necessary to take into account the age-related characteristics of the child’s body. Thus, in children aged 2-12 years, criteria can be used to assess ventricular hypertrophy.
| Left ventricular hypertrophy | Right ventricular hypertrophy | ||
| 1. | R aVL > 8 mV | 1. | R aVR > 4 mV |
| 2. | R V6 > 25 mV | 2. | RV4 > 17 mV |
| 3. | Q V5-6 > 4 mV at R > 20 mV | 3. | RV1 + S V5 > 9 mV |
| 4. | Internal deviation time (IDT) in V5 > 0.045 s | 4. | R/SV1 > 4 mV |
| 5. | EOS deviation to the left | 5. | Shape of the ventricular complex as qR in V1 |
| 6. | T V5-6 flattened or negative | 6. | VBO in V1 > 0.03 s |
| 7. | EOS deviation to the right > 110o |
With myocardial hypertrophy of both ventricles, diagnosis can be difficult due to the fact that the signs of hypertrophy of both ventricles can be mutually leveled.
M.K. Oskolkova (1986) indicates that in the presence of signs of right ventricular hypertrophy, the following criteria indicate hypertrophy of both ventricles:
- dominant RV5 wave (height may be normal) with high positive T waves in these leads;
- increase in q wave amplitude in V5 and V6;
- EOS deviation to the left.
In the presence of signs of left ventricular hypertrophy, the detection of one of the following criteria indicates hypertrophy of both ventricles:
- dominant R or R' waves in lead V1, and its amplitude may be normal;
- the amplitude of the R wave is greater than the amplitude of the q wave in lead aVR;
- The S wave is larger than the R wave in lead V6.
Disturbances in repolarization processes are affected by the state of the autonomic nervous system. Thus, disturbances in the process of repolarization of the ventricular myocardium at rest are detected in children with hypersympathicotonic autonomic reactivity and initial eutonia (in 28%).
During functional tests, disturbances in the process of repolarization of the ventricular myocardium are detected in 25% of children with hypersympathicotonic and in 50% of children with asympathicotonic autonomic reactivity, regardless of the initial autonomic tone.
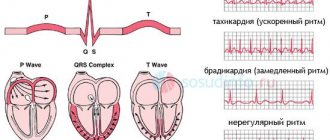
The severity of respiratory arrhythmia is one of the important indicators of the functional state of the heart: if fluctuations in the duration of the R-R intervals exceed 0.3 s, sinus arrhythmia indicates a violation of the regulation of the sinus node and may be a sign of overtraining.
It is important to identify extrasystole. Along with extrasystole at rest, extrasystole can be observed directly during work and in the recovery period. It is generally accepted that extrasystolic arrhythmia recorded during physical activity indicates a certain prepathological state of the heart muscle. Extrasystole at rest is considered more favorable. Some authors believe that the disappearance of extrasystole recorded at rest during physical activity indicates the “harmless” nature of the latter. However, it should be noted that recent experience in sports medicine, obtained through long-term radiotelemetric observations, indicates that extrasystole is an unfavorable sign, regardless of whether it is recorded at rest or during physical activity.
The causes of extrasystoles in athletes are very diverse. They can develop as a result of myocardial overstrain, in diseases resulting in intoxication of the heart muscle, in violation of the nervous regulation of cardiac activity, disorders of mineral metabolism, etc. It must be borne in mind that in athletes, during muscular work, the content of catecholamines in the blood naturally increases, increasing the excitability of the myocardium.
Subjectively, extrasystole in athletes is usually felt in the form of “interruptions” in the work of the heart. With such sensations, an electrocardiographic study is necessary. The occurrence of extrasystoles in athletes should be analyzed by the coach and the doctor in the light of the individual training regimen.
First-degree atrioventricular block (P-Q interval greater than 0.22 s) is observed in athletes with severe fatigue or overtraining and requires medical intervention and significant correction of the training regimen.
Intraventricular conductivity in athletes is most often at the upper limit of normal, which corresponds to the range of 0.06-0.09 s. In some athletes (especially with myocardial hypertrophy), the duration of the QRS complex may be equal to 0.1 s.
When WPW syndrome is detected, careful monitoring is necessary. Classes are often prohibited due to the development of a dangerous rhythm disorder - paroxysmal tachycardia, which should not be underestimated.
Athletes often have first-degree or second-degree Mobitz type I atrioventricular block, which primarily reflects increased vagal tone. An increase in P wave voltage and QRS complex in combination with T wave inversion in the lateral leads may also be detected. QRS prolongation, deviation of the electrical axis of the heart, supraventricular and ventricular tachycardia are not characteristic signs and may require further research.
Many athletes experience a decrease in the voltage of T-waves in the standard and left chest leads, and also have low rates of relative metabolic support of the myocardium. These changes are especially pronounced during accelerated training in adolescence.
Detailed data on ECGs for athletes are given in the book JI.A. Butchenko et al. (1980). According to their data, the electrical axis of the heart (EOS) is normal in 50% of athletes, vertical in 30%, deviated to the right in 4%, horizontal in 12%, and deviated to the left in 3%. Vertical EOS is more often observed in football players, skiers, swimmers, water polo players and asthenics under 20 years of age. Horizontal EOS and deviated to the left are more typical for wrestlers, weightlifters, long and middle distance runners and hypersthenics aged 30-49 years.
P-Q is longer in athletes developing the quality of endurance. A number of features concern the teeth of intervals and ECG segments.
HYPERTROPHY
HYPERTROPHY (from hyper... and Greek τροφή - nutrition), an increase in the volume of an organ, tissue, cells, intracellular structures. It occurs both normally and in various diseases, ensuring the stability of homeostasis. G., along with an increase in volume, is often based on an increase in the number of structural units (organ, tissue, cell), that is, hyperplasia. For example, the G. of a parenchymal organ is based on the G. of parenchymal cells, but at the same time the number of stromal elements can increase, and the formation of new vessels (neoangiogenesis) occurs, providing an increased need for nutrition of hypertrophies. organ. Cell growth also occurs due to an increase not only in the volume of its structures, but also in their number. Thus, G. and hyperplasia are two inextricably linked processes.
Normally, G. develops with increased functional demand or neurohormonal stimulation (for example, myocardial G. in athletes, an increase in the volume of the uterus and mammary glands during pregnancy).
Pathological G. can be true (an increase in the mass and functional activity of an organ due to the G. of cells providing this activity) and false (due to the growth of connective tissue of the stroma and adipose tissue with possible atrophy of parenchymal cells).
True G. is divided into working (compensatory), vicarious (replacement), regenerative, neurohumoral G. and hypertrophic. overgrowth.
Working G. develops with increased work of an organ and is characterized by an increase in the volume of its functionally active structures. For example, the cause of myocardial gastrointestinal tract may be various. hemodynamic factors - arterial and pulmonary hypertension, congenital or acquired heart defects. The weight of the heart can exceed normal values by 3–4 times. G. of the bladder wall develops in men with prostatic hyperplasia, which narrows the urethra and secretes hormonal and growth factors that stimulate G. of detrusor cells (the muscle that expels urine).
Vicarious G. develops in one of the paired organs (lung, kidney, adrenal gland, etc.) following the death or surgical removal of the other.
Regenerative blood forms in the parenchymal cells of the myocardium, neurons, hepatocytes, and others around scars during incomplete regeneration.
With neurohumoral G. and hypertrophic. growths, in fact, we are talking about an increase in the volume of the organ basically. due to the processes of hyperplasia of cellular structures, which simultaneously increase in number and volume.
Molecular mechanisms of cardiac hypertrophy: Part 1. Physiological hypertrophy
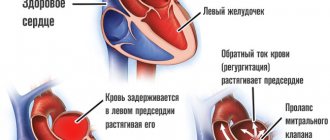
Cardiac hypertrophy is a compensatory reaction that occurs in response to an increase in the load on the myocardium. Physiological hypertrophy leads to increased myocardial function: increased ejection fraction, increased contractile activity.
There are two types of cardiac hypertrophy: physiological and pathological, which differ in molecular mechanisms, cardiac phenotype and prognosis for life. The latter is the most important in clinical practice, since physiological hypertrophy helps to “push” blood through at higher loads, while pathological hypertrophy is associated with unfavorable conditions, including myocardial infarction, arrhythmias, and in the long term can lead to death. It should also be noted that these hypertrophy variants are “competitive antagonists”; There is a constant “struggle” between various signaling systems, the result of which will be the development of either physiological or pathological hypertrophy. [1].
With physiological hypertrophy, there is a slight increase in heart weight (by 10–20%), an increase in cardiomyocytes (CMC) and their proliferation, while the ejection fraction is preserved or increased. There are no sclerotic or necrotic changes in the tissue, and most importantly, physiological hypertrophy is completely reversible (with the exception of postnatal hypertrophy) [1].
Mechanisms of physiological hypertrophy
The initial stimuli leading to hypertrophy do not differ in the case of physiological or pathological variants (for example, physical activity or arterial hypertension). But the reactions triggered in this case are radically different. For the development of physiological hypertrophy, the following is required: an increase in cell size, increased mitochondrial function and energy production, angiogenesis proportional to cell growth, the work of antioxidant systems, regulation of proliferation and regeneration of cardiomyocytes [1].
Physiological hypertrophy of the heart is observed from the birth of a child until he grows up (also called postnatal hypertrophy), with increased physical activity, as well as during pregnancy [2].
Mechanosensors
Mechanotransduction is a fundamental process, the essence of which is the transformation of mechanical signals into biochemical ones. There are sensory systems in the heart that perceive mechanical signals (changes in pressure, volume, etc.) and activate signaling systems responsible for physiological hypertrophy [2].
Ion channels with transient receptor potential (TRP) are one such system. TRPs are a superfamily of transmembrane proteins that function as nonselective ion channels. Among them there are 7 subfamilies; these receptors differ in localization and their role in various cellular processes [3].
In addition to TRP, various integrins exhibit mechanosensitivity. These are transmembrane proteins that can interact with the extracellular matrix (and its various components: fibronectin, laminin, collagen). In response to an extracellular agent, integrins can transmit information into the cell through special integrin-associated protein complexes. This may be focal adhesion kinase (FAK - focal adhesion kinase), integrin-linked kinase (ILK - integrin-linked kinase). The latter, in turn, activate various signaling pathways—standard protein kinase C or PI3K/Act (see below) [2].
We should dwell in somewhat more detail on the integrin-associated protein specific to muscle tissue—melusin. It interacts with β1-integrins, promoting the development of concentric hypertrophy while maintaining cardiac contractility. An increase in melusin synthesis initiates physiological hypertrophy and prevents its development along a pathological path. The opposite is also true: a lack of melusin leads to the development of dilated cardiomyopathy [1].
And, of course, one cannot fail to mention the role of the Z-line as a mechanoreceptor. This part of the cardiomyocyte also responds to stretch by transmitting a signal to many different proteins: teletonin, myopalladin, ankyrins, obscurin... Ultimately, all these processes turn out to be essential for the development of physiological hypertrophy, while defects in some of these proteins lead to cardiomyopathies [2] .
Insulin and insulin-like growth factor 1
These substances regulate many different cellular processes in the heart: proliferation, differentiation, cell growth, metabolism, contractility and apoptosis. Insulin resistance is common in heart failure. IGF1 (insulin-like growth factor 1) is structurally identical to insulin; this agent is synthesized in the liver in response to GH, but can also be formed in other organs (including the heart) [1].
Insulin is known to bind to the insulin receptor, tyrosine kinase, which phosphorylates insulin receptor substrates 1 and 2 (IRS1, 2-insulin receptor substrate). IRSs in turn activate the PI3K/Act signaling pathway. This signaling pathway may be familiar to the reader as a potential target for antitumor therapy. PI3K/Act is involved in the regulation of the cell cycle, and in our case ensures the growth of cardiomyocytes in response to increased load on the myocardium [2], [5].
IGF1, in turn, binds to a specific receptor IGF1R (IGF1 receptor) with subsequent activation of a number of signaling pathways: RAS–RAF–MEK–MAPK, PLC–IP3R3 and others. This also results in physiological hypertrophy of cardiomyocytes.
A little more detail should be taken on the PI3K/Act pathway (it is noteworthy that this signaling pathway is activated during physical activity). PI3K is an enzyme kinase that catalyzes phosphoinositol 3,4,5-trisphosphate. Activation of one of its catalytic subunits—p110α—initiates the processes of physiological hypertrophy and prevents pathological hypertrophy (as we remember, these processes compete with each other). ACT1 is another kinase that is activated by phosphoinositol 3,4,5-phosphate and results in changes in cell metabolism. This is achieved by inhibiting a number of enzymes: glycogen synthase kinase 3β (this kinase inhibits translation initiation factors), fox-O3 (a factor that inhibits the general metabolism of proteins in the cell) and others. As a result, active protein synthesis occurs and the cell increases in size [1].
Triiodothyronine
T3 is triiodothyronine, a thyroid hormone that influences both postnatal cardiac hypertrophy and exercise-induced hypertrophy (although the latter is less reliable) [2].
Immediately after birth, the concentration of T3 in the blood increases significantly. The hormone binds to TRα and TRβ receptors (thyroid hormone receptors), which leads to a “switch” of transcription of the MYH7 gene to MYH6 (these genes encode sequences of myosin heavy chains). MYH7 encodes the β-heavy chains of myosin, MYH6 encodes the α-isoform [1]. This is extremely important, since the α-isoform has greater ATPase activity and, as a result, a cell with active MYH6 is able to contract more strongly [2], [6].
The interaction of T3 with retinoic acid receptors (nuclear receptors for steroid and thyroid hormones, approx.) leads to an increase in the expression of calcium ATPase-2 (SERCA2 - sarcoplasmatic/endoplasmatic reticular calcium ATPase-2), inhibiting MYH7 and the expression of phospholamban (a protein in cardiomyocytes that suppresses SERCA2) [2].
SERCA2 is the evolutionarily most ancient isoform of mammalian calcium ATPases. There are three such isoforms (with subtypes). SERCA2 has three subtypes: a, b and c; for the heart, SERCA2a is most specific (97.5%), there is also a small amount of SERCA2b (2.5%). The function of this protein is to control cytosolic Ca2+ and, as a consequence, to regulate the contractile function of the entire cardiomyocyte [7].
Phospholamban, whose work is inhibited by T3, is a protein regulator of SERCA2 (more precisely, the most studied of these regulators). In its active - dephosphorylated - state, phospholamban suppresses SERCA2 and leads to a decrease in the concentration of Ca2+ in the cytoplasm, which, in turn, reduces the contractility of the cardiomyocyte [2], [7].
In addition, T3 also activates the transcription of β1-adrenergic receptors, sodium and calcium channel proteins, cardiac troponin I, sodium/calcium pump proteins, and adenylate cyclase types V and VI [1], [2]. The hormone also increases the content of TRα1, which activates the already known PI3K [2].
The end result of the work of triiodothyronine in cardiomyocytes is increased myocardial contractile activity and cardioprotection [1].
Nitric oxide (NO)
Physical activity stimulates β3-adrenergic receptors in endothelial cells; in response, endothelial NO synthase (NOS3) is activated, which synthesizes nitric oxide itself, a known vasoactive substance. After this, the following happens:
NO activates soluble guanylate cyclase → cGMP content increases → cGMP-dependent protein kinase G (PKG) is activated → PKG phosphorylates two regulatory proteins (RGS2 and RGS4), which inhibit G-coupled receptors responsible for the development of pathological hypertrophy.
This scheme is needed so that hypertrophy develops according to the physiological variant. A defect in either β3-adrenergic receptors, or NOS3, or the RGS2 and RGS4 proteins leads to a break in the entire chain of cardioprotection, which may well result in myocardial infarction [1].
Angiogenesis
Cells need sufficient blood supply to function normally. If vascular density increases in proportion to the growth of cardiomyocytes, physiological hypertrophy develops. But when cells grow faster than the capillary network can allow, chronic hypoxia occurs.
One of the most important factors determining angiogenesis is vascular endothelial growth factor (VEGF). VEGF may also be familiar to the reader - inhibitors of this factor are used in the treatment of various types of cancer. However, such drugs have significant cardiotoxicity and can lead to various cardiomyopathies (which indirectly indicates the importance of VEGF for myocardial function). Among other things, changes in VEGF levels are also associated with peripartum (postpartum) cardiomyopathy [1], [2].
At the beginning of the article, NFAT signaling molecules were mentioned (in the “mechanosensors” section). VEGF, influencing endothelial cells, acts precisely through the NFAT system, causing enhanced angiogenesis. Thus, these nuclear factors, in addition to their anti-apoptotic role, also affect the blood supply to cardiomyocytes [8].
In addition to VEGF, platelet-derived growth factor (PDGD) also plays an important role. The mechanisms by which PDGD influences angiogenesis are not known for certain; however, a defect in this factor in experiments on mice leads to heart failure and deterioration of blood supply [1].
It is also impossible not to mention another important substance. Hypoxia-inducible factor 1α (HIF1α - hypoxia-inducible factor) is a major transcription factor that ensures a constant supply of oxygen to cells by regulating angiogenesis, vascular modification and regulation of glucose metabolism. In the case of pathological hypertrophy, the p53 antigen (known for its association with tumor development) activates the ubiquitination and proteasomal degradation of HIF1α [1]. Normally, HIF1α activates various signaling molecules (including VEGF) and promotes enhanced angiogenesis.
Neuregulin-1
Neuregulins (1-4) are members of the epidermal growth factor family. In the cardiovascular system, neuregulin-1 is mainly present. The signaling pathway of this factor (and its receptors) plays an important role in the adaptation of the myocardium to the load and the formation of the heart muscle.
Neuregulin-1 acts on the ErbB family kinase (a group of molecules structurally similar to epidermal growth factor; proteins of this family are often considered as potential tumor markers, approx.). Neuregulin-1 activates ErbB2 and ErbB4, which leads to stimulation of the already mentioned PI3K system. The result of this interaction is the proliferation of cardiomyocytes, which prevents ischemic lesions [1].
MicroRNAs and RNA-binding proteins
MicroRNAs (miRNAs) are expressed in the heart during physiological hypertrophy induced by aerobic exercise. For example, miRNA-222 in mice inhibits four targets potentially responsible for decompensation and the development of heart failure: p-27 (encodes a cell cycle inhibitor), Hipk1 and Hipk2 (encodes protein kinases), Hmbox1 (encodes a transcription inhibitor). In other words, activation of miRNA-222 promotes the growth and proliferation of cardiomyocytes [1].
Also, any form of hypertrophy requires de novo protein synthesis. And here, eukaryotic translation factor 4F (elF4F) and the mammalian target of rapamycin mTORC1 are involved - together they trigger increased protein synthesis. In addition, in response to a physiological (and pathological) stimulus in myocardial cells, the poly-A sequence elongates and the expression of specific polyadenylate-binding protein 1 (PABPC1 - polyadenylate-binding protein 1) occurs, which binds to elF4F and thus promotes accumulation in cells various construction (and other) proteins. Increased expression of PABPC1 in cardiomyocytes is necessary for physiological hypertrophy (nothing is known yet about the role of the protein in its pathological version) [1].
Sources:
- M. Nakamura and J. Sadoshima, 'Mechanisms of physiological and pathological cardiac hypertrophy', Nat. Rev. Cardiol., vol. 15, no. 7, pp. 387–407, 2021.
- M. Maillet, J. H. Van Berlo, and J. D. Molkentin, 'Molecular basis of physiological heart growth: Fundamental concepts and new players', Nat. Rev. Mol. Cell Biol., vol. 14, no. 1, pp. 38–48, 2013.
- B. Nilius and G. Owsianik, 'The transient receptor potential family of ion channels', Genome Biol., vol. 12, no. 3, 2011.
- W. T. Pu, Q. Ma, and S. Izumo, 'NFAT transcription factors are critical survival factors that inhibit cardiomyocyte apoptosis during phenylephrine stimulation in vitro', Circ. Res., vol. 92, no. 7, pp. 725–731, 2003.
- M. Osaki, M. Oshimura, and H. Ito, 'PI3K-Akt pathway: Its functions and alterations in human cancer', Apoptosis, vol. 9, no. 6, pp. 667–676, 2004.
- E. M. McNally, R. Kraft, M. Bravo-Zehnder, D. A. Taylor, and L. A. Leinwand, 'Full-length rat alpha and beta cardiac myosin heavy chain sequences', J. Mol. Biol., vol. 210, no. 3, pp. 665–671, 1989.
- P. Vangheluwe, L. Raeymaekers, L. Dode, and F. Wuytack, 'Modulating sarco(endo)plasmic reticulum Ca2+ ATPase 2 (SERCA2) activity: Cell biological implications', Cell Calcium, vol. 38, no. 3-4 SPEC. ISS., pp. 291–302, 2005.
- T. A. Zaichuk, E. H. Shroff, R. Emmanuel, S. Filleur, T. Nelius, and O. V. Volpert, 'Nuclear factor of activated T cells balances angiogenesis activation and inhibition', J. Exp. Med., vol. 199, no. 11, pp. 1513–1522, 2004.