Action potential (AP) is a short-term amplitude change in the resting membrane potential (RMP) that occurs when a living cell is excited. Essentially, this is an electrical discharge - a rapid, short-term change in potential in a small area of the membrane of an excitable cell (neuron or muscle fiber), as a result of which the outer surface of this area becomes negatively charged in relation to neighboring areas of the membrane, while its inner surface becomes positively charged in relation to to adjacent areas of the membrane. The action potential is the physical basis of a nerve or muscle impulse that plays a signaling (regulatory) role.
general characteristics
Action potentials can differ in their parameters depending on the type of cell and even on different parts of the membrane of the same cell. The most typical example of differences is the action potential of the heart muscle and the action potential of most neurons. Nevertheless, the basis of any action potential is the following phenomena:
- “The membrane of a living cell is polarized” - its inner surface is charged negatively in relation to the outer one due to the fact that in the solution at its outer surface there is a larger number of positively charged particles (cations), and at the inner surface there is a larger number of negatively charged particles (anions).
- “A membrane has selective permeability” - its permeability to different particles (atoms or molecules) depends on their size, electrical charge and chemical properties.
- “The membrane of an excitable cell is capable of quickly changing its permeability for a certain type of cations, causing a transition of positive charge from the outside to the inside
The first two properties are characteristic of all living cells. The third is a feature of excitable tissue cells and the reason why their membranes are able to generate and conduct action potentials.
The main mathematical model describing the generation and transmission of action potentials is the Hodgkin-Huxley model.
Phases
Five phases of development of PD can be clearly distinguished:
Rising (depolarization)
The occurrence of an action potential (AP) is associated with an increase in the permeability of the membrane for sodium ions (20 times compared to the permeability for K +, and 500 times compared to the initial permeability of Na +) and a subsequent increase in the diffusion of these ions along the concentration gradient into the cell, leads to a change (decrease) in membrane potential.
A decrease in membrane potential leads to an increase in membrane permeability to sodium by opening voltage-gated sodium channels, and an increase in permeability is accompanied by increased diffusion of sodium into the cytoplasm, which causes even more significant depolarization of the membrane. Due to the presence of positive feedback, depolarization of the membrane during excitation occurs with acceleration and the flow of sodium ions into the cell increases all the time. The intensity of the flow of potassium ions directed from the cell outwards in the first moments of excitation remains at the beginning. The increased flow of positively charged sodium ions into the cell first causes the disappearance of excess negative charge on the inner surface of the membrane, and then leads to recharging of the membrane. The influx of sodium ions occurs until the inner surface of the membrane acquires a positive charge sufficient to balance the sodium concentration gradient and stop its further passage into the cell. The sodium occurrence of PD is confirmed by experiments with changes in the external and internal concentrations of this ion. It was shown that a tenfold change in the concentration of sodium ions in the external or internal environment of the cell corresponds to a change in PD of 58 mV. When sodium ions were completely removed from the fluid surrounding the cell, PD did not occur. Thus, it has been established that AP occurs as a result of excess, compared to rest, diffusion of sodium ions from the surrounding fluid into the cell. The period during which the membrane permeability for sodium ions increases when sodium channels open is short (0.5-1 ms), followed by an increase in membrane permeability for potassium ions due to the opening of voltage-dependent potassium channels, and, consequently, increased diffusion these ions out of the cell. The “all or nothing” principle According to the “all-or-nothing” law, the cell membrane of an excitable tissue either does not respond to the stimulus at all, or responds with the maximum force possible for it at the moment. The action of the stimulus usually leads to local depolarization of the membrane. This causes the opening of sodium channels, which are sensitive to changes in potential, and through this increases sodium conductance, which leads to even greater depolarization. The existence of such feedback ensures regenerative (renewable) depolarization of the cell membrane. The magnitude of the action potential depends on the strength of the stimulus, and it occurs only when depolarization exceeds a certain limiting level specific for each cell. This phenomenon is called “all or nothing.” However, if depolarization is 50-75% of the limiting value, then a local response may occur in the cell, the amplitude of which is significantly lower than the amplitude of the action potential. The absence of an action potential at the pidboundary level of depolarization is explained by the fact that sodium permeability does not increase sufficiently to cause regenerative depolarization. The level of depolarization that occurs does not cause the opening of new sodium channels, so sodium conductance quickly decreases, and the resting potential in the cell is again established.
Overshoot
Depolarization of the membrane leads to a reversal of the membrane potential (the MP becomes positive).
In the overshoot phase, the Na + current begins to rapidly decrease, which is associated with the inactivation of voltage-dependent Na + channels (the open state time is milliseconds) and the disappearance of the electrochemical Na + gradient. Refractoriness One of the consequences of the disappearance of the Na + gradient is refractoriness - a temporary inability to respond to a stimulus. If the stimulus occurs immediately after the passage of the action potential, then excitability will not occur either with a stimulus strength at the threshold level or with a significantly stronger stimulus. This state of complete inexcitability is called the absolute refractory period. This is followed by a relative refractory period, when a suprathreshold stimulus can cause an action potential with a significantly lower amplitude than normal. An action potential of the usual amplitude under the action of a threshold stimulus can be evoked only after a few milliseconds after the preliminary action potential. The absolute refractory period limits the maximum frequency of generation of action potentials.
Repolarization
An increase in the potassium ion flow directed outward from the cell leads to a decrease in the membrane potential, which in turn causes a decrease in the permeability of the membrane to sodium ions, which, as indicated, is a function of the membrane potential. Thus, the second stage is characterized by the fact that the flow of potassium ions from the cell outward increases, and the counter flow of sodium ions decreases. This membrane repolarization continues until the resting potential is restored—membrane repolarization. After this, the permeability to potassium ions also drops to its original value. Due to the positively charged potassium ions released into the environment, the outer surface of the membrane again acquires a positive potential relative to the internal one.
Trace depolarization and hyperpolarization
In the final phase, the restoration of the resting membrane potential slows down, and trace reactions are recorded in the form of trace depolarization and hyperpolarization, due to the slow restoration of the initial permeability for K + ions.
Spontaneous diastolic depolarization and automaticity
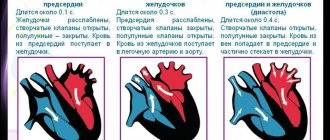
The membrane potential of normal cells of the working myocardium of the atria and ventricles remains constant at the level of the resting potential throughout diastole (see Fig. 3.1): if these cells are not excited by a propagating impulse, then the resting potential in them is maintained indefinitely. In other types of cardiac fibers, such as the specialized fibers of the atria or the Purkinje fibers of the ventricular conduction system, the membrane potential is not constant during diastole and gradually changes towards depolarization.
If such a fiber is not excited by a propagating impulse before the membrane potential reaches a threshold level, then a spontaneous action potential may arise in it (Fig. 3.6). The change in membrane potential during diastole is called spontaneous diastolic depolarization, or phase 4 depolarization.
Causing the occurrence of action potentials, this mechanism serves as the basis for automatism. Automaticity is a normal property of the cells of the sinus node, the muscle fibers of the mitral and tricuspid valves, some parts of the atria, the distal part of the AV node, as well as tissues of the His-Purkinje system.
In a healthy heart, the rate of impulse generation due to the automaticity of sinus node cells is high enough to allow propagating impulses to excite other potentially automatic cells before they spontaneously depolarize to a threshold level. In this case, the potential automatic activity of other cells is usually suppressed, although it can manifest itself in a number of physiological and pathological conditions (discussed below).
Rice. 3.6. Spontaneous diastolic depolarization and automaticity of Purkinje fibers in a dog. A - spontaneous excitation of Purkinje fibers at a maximum diastolic potential of -85 mV. Diastolic depolarization is a consequence of a decrease in time of the current ins, or pacemaker current (see text). B - automatic activity that occurs when the membrane potential decreases;
with a larger pulse amplitude, the membrane potential decreases to -40 mV, causing maintained rhythmic activity. Fragment B3: an increased current pulse reduces the membrane potential to -30 mV, as a result of which maintained rhythmic activity occurs with greater frequency. Such rhythmic activity, which occurs at potentials less negative than -60 mV, probably depends on a different pacemaker current than the activity indicated on fragment A.
Spontaneous diastolic depolarization is a consequence of a gradual change in the balance between incoming and outgoing membrane currents in favor of the total incoming (depolarizing) current. When studying the pacemaker current using the voltage clamp method in Purkinje fibers [34–37] and ganglion cells [38], the dependence of the characteristics of the portal system on both potential and time was shown.
Based on initial studies of the potential level at which the pacemaker current reverses its direction, it was assumed that the outward pacemaker current, carried by K ions, is gradually deflected, thereby allowing the inward background current to depolarize the cell membrane [34–36].
However, according to the interpretation of the results of more recent experiments, the normal pacemaker current is an inward current carried predominantly by Na ions, which increases over time, thereby causing gradual diastolic depolarization [37, 38]. When depolarization reaches the threshold potential level, an impulse occurs, after which pacemaker conductance is inactivated during membrane depolarization and can only be reactivated after repolarization of the action potential.
It is clear that the frequency of spontaneous excitations is determined by the time during which diastolic depolarization changes the membrane potential to the threshold level; therefore, changes in threshold potential or rate of diastolic depolarization, such as those occurring in Purkinje fibers under the influence of adrenaline, may affect the frequency of automatic activity.
In the cells of the sinus and atrioventricular nodes, the rate of depolarization in the zero phase is much lower (1-20 V/s) than in normal Purkinje fibers or cells of the working myocardium (see Fig. 3.9). The amplitude of action potentials is also very small (60-80 mV); in some cells the peak of the action potential does not exceed 0 mV [3].
Compared with other cardiac cells, the lower rise rate and smaller amplitude of the nodal cell action potential reflects the significantly lower magnitude of inward current before the zero depolarization phase in these cells. Currently available data clearly indicate that the smaller incoming current in the cells of the sinus and atrioventricular nodes does not flow through fast sodium channels, but through slow channels and is carried by sodium and calcium ions [45-47].
Such action potentials with a rise dependent on a slow inward current are often called “slow responses” in contrast to the more common “fast responses” whose rise depends on a fast sodium current [20]. Due to such a small value of the total incoming current and slow depolarization in the zero phase, the speed of slow action potentials through the nodes is always low (0.01-0.1 m/s);
It is precisely this slow conduction that, under certain conditions, can cause rhythm disturbances in the tissues of the nodes. As noted earlier, slow inward current channels have completely different time and potential gating characteristics compared to fast sodium channels.
The slow incoming current takes much longer to activate and inactivate than the fast sodium current. Therefore, after the action potential rises in the nodes, the slow inward current is inactivated only slowly, promoting membrane depolarization throughout the plateau phase of the action potential. Activation of the time- and potential-dependent outward potassium current, together with inactivation of the slow inward current, appears to induce repolarization of nodal cells, as has been described for action potential termination in other cardiac cells.
Rice. 3.9. Comparison of the action potentials of the sinus and AV node (indicated by arrows) with the action potentials of the working myocardium and Purkinje fibers. Action potentials were recorded in the following areas of the heart (starting from the top): sinus node, atrium, atrioventricular node, His bundle, Purkinje fiber in the false tendon , terminal Purkinje fiber and working ventricular myocardium.
Note that the increase in velocity and amplitude of action potentials in the sinus node and AV node is less than those in other cells [3]. The conductance of slow channels for inward current is also restored after membrane repolarization much more slowly than the conductance of fast sodium channels [20, 21].
Unlike other cardiac cells, in nodal cells, when a premature stimulus is applied during the final phase of repolarization, no action potential is observed. In fact, sufficient inactivation of the conductance of slow channels for inward current can persist even after complete repolarization in cells that have become completely refractory to stimulation [48].
Reactivation occurs gradually throughout diastole; premature impulses, caused soon after complete repolarization, have a slower rise, lower amplitude than normal impulses, and propagate more slowly. Premature impulses evoked later in diastole have a correspondingly faster rise time, higher amplitude, and are therefore more quickly conducted [49].
This behavior reflects the duration of the reactivation process of slow channels. The associated long refractory period of the nodal tissue, as well as the significant slowdown in the conduction of premature impulses through it, may be important factors in the initiation of some cardiac arrhythmias. Automaticity The cells of the sinus node are usually automatic, so that each subsequent action potential arises as a result of spontaneous diastolic depolarization;
AV node cells are also capable of firing automatically, especially if they are not connected to the surrounding atrial myocardium [50]. This means, apparently, that the electrotonic interaction between the cells of the node and the atria is suppressed automatically through the atrionodal conduction. The automatic activity of sinus node cells cannot be associated with the same pacemaker current that was previously described in Purkinje fibers.
Changes in the state of the membrane conductance gate mechanism, which is responsible for the normal automaticity of Purkinje fibers, occur only in the membrane potential range from -90 to -60 mV [36]. Such conduction changes are unlikely to explain the development of spontaneous diastolic depolarization in sinus node cells, since the maximum diastolic potential of these cells is usually less negative than -60 mV.
However, available evidence suggests that the pacemaker current in the sinus node is carried, at least in part, by K ions [38] and a decrease in this outward current in the presence of a constant background inward current leads to a gradual depolarization of the membrane. In addition, an important role is probably played by the inward current, which is activated by hyperpolarization and is designated if [38].
https://infopedia.su/12x90c6.html
The electrical impulse that travels through the heart and triggers each contraction cycle is called an action potential; it represents a wave of short-term depolarization, during which the intracellular potential in each cell in turn becomes positive for a short time and then returns to its original negative level.
Changes in the normal cardiac action potential have a characteristic development over time, which for convenience is divided into the following phases [3]: phase 0 - initial rapid depolarization of the membrane; phase 1 - rapid but incomplete repolarization; phase 2 - “plateau”, or prolonged depolarization, characteristic of the action potential of cardiac cells;
phase 3 - final fast repolarization; phase 4 - diastole period. During the action potential, the intracellular potential becomes positive, since the excited membrane temporarily becomes more permeable to Na (compared to K), so the membrane potential for some time approaches the equilibrium potential of sodium ions (ENa) - EN can be determined using the Nernst relation;
at extracellular and intracellular Na concentrations of 150 and 10 mM, respectively, it will be: However, the increased permeability for Na persists only for a short time, so that the membrane potential does not reach ENa and after the end of the action potential returns to the resting level. The above changes in permeability, causing the development of the depolarization phase of the potential actions arise due to the opening and closing of special membrane channels, or pores, through which sodium ions easily pass.
The gating is believed to regulate the opening and closing of individual channels, which can exist in at least three conformations—open, closed, and inactivated. One gate, corresponding to the activation variable "m" in the Hodgkin-Huxley description [13] of sodium ion currents in the membrane of the squid giant axon, moves rapidly to open the channel when the membrane is suddenly depolarized by a stimulus.
The other gate, corresponding to the inactivation variable “h” in the Hodgkin-Huxley description, moves more slowly during depolarization, and its function is to close the channel (Fig. 3.3). Both the steady-state distribution of gates within the channel system and the rate of their transition from one position to another depend on the level of membrane potential.
Therefore, the terms "time-dependent" and "voltage-dependent" are used to describe membrane Na conductance. If a resting membrane is suddenly depolarized to a positive potential (for example, in a voltage clamp experiment), the activation gate will quickly change its position to open sodium channels , and then the inactivation gate will slowly close them (Fig. 3.3).
The word "slow" here means that inactivation takes a few milliseconds, while activation occurs in a fraction of a millisecond. The gates remain in these positions until the membrane potential changes again, and for all gates to return to their original resting state, the membrane must be completely repolarized to a high negative potential level.
If the membrane is repolarized only to a low level of negative potential, then some inactivation gates will remain closed and the maximum number of available sodium channels that can open upon subsequent depolarization will be reduced [14]. (The electrical activity of cardiac cells in which sodium channels are completely inactivated will be discussed below.
Spreading
Spread into unmyelinated fibers
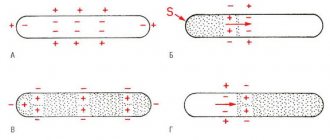
In unmyelinated (without pulp) nerve fibers, the AP spreads from point to point, since excitation can be registered as one that gradually “runs” along the entire fiber from its point of origin. Sodium ions entering the excited area serve as a source of electric current for the occurrence of AP in adjacent areas. In this case, the impulse occurs between the depolarized section of the membrane and its non-excited section. The potential difference here is many times higher than necessary for membrane depolarization to reach the maximum level. The speed of pulse propagation in such fibers is 0.5-2 m/s
Spread in myelinated fibers
The nerve processes of most somatic nerves are myelinated.
Only very small areas of them, the so-called node interception (interception of Ranvier), are covered with a normal cell membrane. Such nerve fibers are characterized by the fact that voltage-dependent ion channels are located on the membrane only at the interceptions. In addition, this shell increases the electrical resistance of the membrane. Therefore, when the membrane potential shifts, the current passes through the membrane of the intercepting area, that is, by jumping (saltatory) from one interception to another, which allows you to increase the speed of the nerve impulse, which ranges from 5 to 120 m/s. Moreover, the action potential that arose in one of the nodes of Ranvier causes action potentials in neighboring nodes due to the emergence of an electric field, which causes an initial depolarization in these nodes. The parameters of the EMF field and the distance of its effective action depend on the cable properties of the axon. Types of nerve fibers, impulse conduction speed, depending on myelination
| Type | Diameter (µm) | Myelination | Conduction speed (m/s) | Functional purpose |
| A alpha | 12-20 | strong | 70-120 | Mobile fibers of the somatic NS; proprioceptor sensory fibers |
| A beta | 5-12 | strong | 30-70 | Sensory fibers of skin receptors |
| A gamma | 3-16 | strong | 15-30 | Sensory fibers of proprioceptors |
| A delta | 2-5 | strong | 12-30 | Sensitive fibers of thermoreceptors, nociceptors |
| IN | 1-3 | weak | 3-15 | Preganglionic fibers of the sympathetic nervous system |
| WITH | 0,3-1,3 | absent | 0,5-2,3 | Postganglionic fibers of the sympathetic nervous system; sensory fibers of thermoreceptors, nociceptors of some mechanoreceptors |
Diagnostics
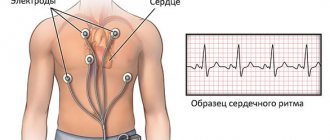
Cardiac electrocardiography (ECG) is used to diagnose the correct processes of depolarization and repolarization.
The teeth and intervals mean nothing to the layman. Functional diagnostic doctors are familiar with subtle signs and changes in characteristic waves and can calculate the time of repolarization.
An increase in the depolarization time of the ventricles of the heart indicates a mechanical obstacle in the propagation of the impulse. This is possible with blockades of varying degrees. Acute infarction most often affects the left ventricle. Here a connective tissue scar is formed, which serves as an obstacle to the impulse. At the conclusion of the ECG, the doctor, in addition to signs of a heart attack, will definitely write about a moderate depolarization disorder.
When deciphering, the shape of the complexes, the height and width of the teeth, the level of the main line, and the duration of the intervals are taken into account
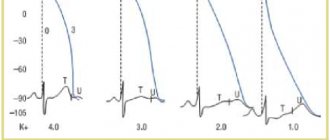
Impaired repolarization is indicated by a decrease in the T wave. This is typical for diffuse dystrophic changes and cardiosclerosis. In this case, the ECG conclusion does not make a diagnosis, but helps to understand the mechanism of formation of symptoms of the disease, stage and form.
Repolarization is disrupted by myocardial hypertrophy, taking certain medications, lack of microelements and vitamins in the diet, and dehydration. Such a patient should be examined in a hospital and a stress test with potassium chloride should be performed. After potassium administration, the ECG shows normalization of the shape of the ventricular complexes.
Action potential propagation between cells
At a chemical synapse, after the action potential wave reaches the nerve terminal, it causes the release of neurotransmitters from the presynaptic vesicles into the synaptic cleft. Transmitter molecules released from the presynapse bind to receptors on the postsynaptic membrane, resulting in the opening of ion channels in the receptor macromolecules. Ions begin to enter the postsynaptic cell through open channels, change the charge of its membrane, which leads to partial depolarization of the membrane and, as a consequence, provoking the generation of an action potential in the postsynaptic cell.
In the electrical synapse there is no “mediator” of transmission in the form of a neurotransmitter. But the cells are connected to each other using specific protein tunnels - conexons, so ionic currents from the presynaptic cell can stimulate the postsynaptic cell, causing the generation of an action potential in it. Thanks to this structure, the action potential can propagate in both directions and much faster than through a chemical synapse.
- Scheme of the process of nerve signal transmission at a chemical synapse
- Diagram of the structure of an electrical synapse
Repolarization disorder
Early ventricular repolarization syndrome
For some reason, there may be a disruption in repolarization processes. This means that there is a problem with charge restoration. The cell does not have time to “rest” before the next excitation. Various reasons may contribute to this. Taking medications that stimulate the sympathetic nervous system can cause the development of early ventricular repolarization syndrome. Connective tissue diseases (collagenosis), cardiomyopathy, heart defects, coronary heart disease, myocardial hypertrophy and other factors can lead to disturbances in repolarization processes.
Clinically, this may manifest itself as symptoms of the underlying disease, or it may be recorded only on an electrocardiogram. However, no matter how things are with your health, you should consult a doctor. Be healthy!
Action potential in different cell types
Action potential in muscle tissue
The action potential in skeletal muscle cells is similar to the action potential in neurons. Their resting potential is typically -90 mV, which is less than the resting potential of typical neurons. The action potential of muscle cells lasts approximately 2–4 ms, the absolute refractory period is approximately 1–3 ms, and the conduction velocity along the muscle is approximately 5 m/s.
Action potential in cardiac tissue
The action potential of the cells of the working myocardium consists of a phase of rapid depolarization, an initial rapid repolarization, which turns into a phase of slow repolarization (plateau phase), and a phase of rapid final repolarization. The phase of rapid depolarization is caused by a sharp increase in the permeability of the membrane for sodium ions, causes a rapid incoming sodium current, when the membrane potential reaches 30-40 mV it is inactivated and subsequently the calcium ion current plays a major role. Depolarization of the membrane causes activation of calcium channels, resulting in an additional depolarizing incoming calcium current.
The action potential in cardiac tissue plays an important role in coordinating cardiac contractions.
Delayed afterdepolarization and triggered sustained rhythmic activity
frequency leads to a decrease in amplitude. In addition, if a premature action potential during stimulation occurs at a constant frequency, then the subsequent afterdepolarization has a greater amplitude than that observed after a regular action potential. Moreover, the earlier during the main cycle the premature action potential occurs, the greater the amplitude of the premature afterdepolarization.
At a high enough frequency of sustained stimulation or after a sufficiently early premature stimulus, afterdepolarization can reach threshold and produce unstimulated action potentials. The first spontaneous impulse is observed after a shorter interval compared to the duration of the main cycle, since the post-depolarization due to which it arose begins soon after the repolarization of the previous action potential.
Consequently, the spontaneous impulse causes another post-depolarization, which also reaches the threshold level, causing the appearance of a second spontaneous impulse (see Fig. 3.8). This last impulse causes the next afterdepolarization, which initiates the third spontaneous impulse, and so on throughout the duration of the trigger activity.
Triggered activity may cease spontaneously and, if it does, the last unstimulated pulse is usually followed by one or more subthreshold afterdepolarizations. Fig. 3.8. Induction of trigger activity in the atrial fiber of the mitral valve in a monkey. Each fragment shows only the lower part of the action potentials.
The horizontal lines in fragments I and II are drawn at a level of -30 mV, and in fragment III - at a level of -20 mV. fragment IA and 1B: trigger activity resulting from a reduction in the duration of the main stimulation cycle. IA: stimulation cycle duration is 3400 ms; and each action potential is followed by a subthreshold delayed afterdepolarization.
in the occurrence of trigger activity due to premature stimulation. IIIA: A premature impulse (arrow) is produced during the repolarization phase of the afterdepolarization, and the amplitude of the subsequent afterdepolarization increases. IIIB: the premature impulse (large arrow) is followed by a post-depolarization that reaches the threshold (small arrow) and leads to the appearance of a series of trigger impulses [40].
Molecular mechanisms of action potential generation
The active properties of the membrane that ensure the occurrence of an action potential, based mainly on the behavior of voltage-gated sodium (Na +) and potassium (K +) channels. The initial phase of AP is formed by the input sodium current, later potassium channels open and the output K + -current returns the membrane potential to the initial level. The initial ion concentration is then restored by the sodium-potassium pump.
During the PD, channels pass from state to state: in Na + channels there are three main states - closed, open and inactivated (in reality everything is more complicated, but these three states are enough for description), in K + channels there are two - closed and open.
The behavior of the channels involved in the formation of PD is described in terms of conductivity and calculated through transfer coefficients.
Carryover coefficients were derived by Alan Lloyd Hodgkin and Andrew Huxley.
Conductivity for potassium GK per unit area
| , |
| Where: |
| an — Transfer coefficient from closed to open state for K+ channels [1/s]; |
| bn — Transfer coefficient from open to closed state for K+ channels [1/s]; |
| n — Fraction of K+ channels in the open state; |
| (1 - n) - Fraction of K + channels in the closed state |
The conductivity for sodium G Na per unit area
is more difficult to calculate, since, as already mentioned, in voltage-dependent Na + channels, in addition to the closed / open states, the transition between which is a parameter, there are also inactivated / not inactivated states, the transition between which is described through a parameter
| , | , |
| Where: | Where: |
| am — Transfer coefficient from closed to open state for Na + channels [1/s]; | ah — Transfer coefficient from inactivated to non-inactivated state for Na + channels [1/s]; |
| bm — Transfer coefficient from open to closed state for Na + channels [1 / s]; | bh — Transfer coefficient from non-inactivated to inactivated state for Na + channels [1 / s]; |
| m — Fraction of Na + channels in the open state; | h — Fraction of Na + channels in a non-inactivated state; |
| (1 - m) - Fraction of Na + channels in the closed state | (1 - h) - Fraction of Na + channels in the inactivated state. |
Story
The main provisions of the membrane theory of excitation were formulated by the German neurophysiologist Yu. Bernstein
In 1902, Julius Bernstein put forward a hypothesis according to which the cell membrane allows K+ ions into the cell, and they accumulate in the cytoplasm. The calculation of the resting potential value using the Nernst equation for the potassium electrode satisfactorily coincided with the measured potential between the muscle sarcoplasm and the environment, which was about -70 mV. According to the theory of Yu. Bernstein, when a cell is excited, its membrane is damaged, and K + ions leave the cell along a concentration gradient until the membrane potential becomes zero. The membrane then restores its integrity and the potential returns to the resting potential level.
This model was developed in their 1952 work by Alan Lloyd Hodgkin and Andrew Huxley, in which they described the electrical mechanisms responsible for the generation and transmission of a nerve signal in the squid giant axon. For this, the authors of the model received the Nobel Prize in Physiology or Medicine for 1963. The model is called the Hodgkin-Huxley model
In 2005, Thomas Heimburg and Andrew D. Jackson proposed the soliton model, based on the assumption that the signal propagates through neurons in the form of solitons - stable waves propagating along the cell membrane.
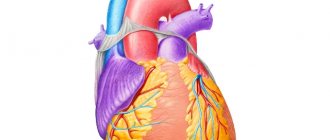
Anatomy of the conduction system
Conduction system of the heart
Quite often, not only the medical worker, but also the patient who comes for an appointment, hears the phrase “repolarization processes.” What is this in essence? Sometimes a lack of knowledge forces the patient to think a lot, and the fruits of reflection lead him to unreasonable worries and sorrows. And all because there is a lack of understanding of the simple truths of electrophysiology. So, our heart, having a conducting system, has the ability to be excited and return to a state of rest. This definition is, of course, conditional. After all, this organ works 24 hours a day, which means that there should be no pauses in the work of a healthy heart.
Let us turn specifically to its electrophysiology. “From head to toe,” our heart is provided with a conducting system, which is represented by nodes, bundles and fibers. The structural unit of this system is the atypical muscle cell. It contains fewer contractile elements of myofibrils, but more sarcoplasm - cytoplasm that fills the space between myofibrils. It is the sarcoplasm that ensures the conduction of exciting impulses inside the fiber. Even externally, these pathways can be distinguished with the naked eye - they are a little paler.
The effect of certain substances on the action potential
Some substances of organic or synthetic origin can block the formation or passage of PD:
- Batrachotoxin has been found in some representatives of the leaf climber genus. Sustainably and irreversibly increases the permeability of membranes to sodium ions.
- Poneratoxin was found in ants of the genus Paraponera. Blocks sodium channels.
- Tetrodotoxin was found in the tissues of fish of the Skelezubovi family, from which the Japanese delicacy Fugu is prepared. Blocks sodium channels.
- The mechanism of action of most anesthetics (Procaine, Lidocaine) is based on blocking sodium channels and, accordingly, blocking the conduction of impulses along sensitive nerve fibers.
- 4-Aminopyridine - reversely blocks potassium channels, prolongs the duration of the action potential. Can be used in the treatment of multiple sclerosis.
- ADWX 1 - reversely blocks potassium channels. Under experimental conditions, it alleviated the course of acute disseminated encephalomyelitis in rats.